


解偶联剂在污泥减量上的应用
0 前言
活性污泥的消化过程是一个利用菌群将有机污染物质转换为细胞内物质和代谢产物的过程。在消化过程中,随着菌群数量的增多生物浓度将急剧增加,从而造成污泥中生物过剩。尽管这种处理处置方法(污泥消化)已经占用了总处理费用的60%,但在欧共体的法规要求下,这些花费将有可能更高,而用垃圾填埋处理则费用将会降低很多。
快速消耗掉菌群合成代谢中的能量而不降低有机污染物的去除率,是一个可以直接降低生物(污泥)产量的有效方法。化学渗透假说中提出的氧化磷酸化理论(在分解代谢过程中产生ATP并放出能量,在合成代谢中消耗能量并产生ATP,反应方程式为:ATP←可逆→ADP+能量)指出:可以通过控制环境中P质子的浓度使能量(ATP)以游离的形式存在,此时生物氧化有机污染物质的反应仍然是是可以继续进行的,但磷酸化时ADP吸收能量转换为ATP的过程却被抑制了,从而限制了生物同化过程所需的能量,生物产量也就随之降低了。
因此,防治生物过剩的目标达到了,同时也因为质子浓度的增加而加速了污染物质与质子的接触机会。这一研究使我们意识到:如果我们在一个只培养有假单细胞的环境里增加P质子(pNP)的浓度,也可以达到相同的处理处置效果。PH值的改变导致质子浓度变化,从而可以诱发生物产量的变化这一现象也同时被发现了。这样一来,费用、安全性、环境保护等因素在污染的处理处置过程中都被考虑到了,并且可以达到最佳的处理效果和平衡状态。
1 实验材料和方法
当在活性污泥中加入一种新的未知的菌群——单一的P-putida NCIMB1005菌落(活性污泥中最重要的要素之一),并将其视为一个简单的模型,用葡萄糖做为唯一的碳源。葡萄糖有限度地连续氧化过程可以通过21个不同稀释比、工作容积为1.51的恒化器(LH发酵器)来维持和控制。温度控制在30℃,pH由一个自动添加1Mol/L硝酸或氢氧化钠的装置控制着。
生物浓度的值可以用紫外分光光度计在波长为600钠米处测量透光度(从而计算出其温度的方法)来测量获得。葡萄糖的浓度可以通过化验-葡萄糖来确定。pNP的浓度可以用酸碱指示剂显示再通过比色获得。
对于恒化器的工作状况,我们提出如下三点假设:
1、微生物都完全悬浮在溶液中。
2、恒化器中的物质是完全混合均匀的,新加入的物质都非常均匀地分散于培养基中,即恒化器是一个理想的完全混合式反应器。
3、培养基一直处于平衡状态(加入量和流出量相平衡),且恒化器的工作容积是恒定的。
培养基的消耗速率可能通过物质平衡原理来描述,即:培养基的总消耗速率=维持生物存活最基本能量需求的消耗速率+生物生长时同化过程对培养基的消耗速率;用公式可以做如下的描述:
-rs=-μX/YG-qmX (1)
其中: rS-培养基的消耗速度(g〔培养基〕/h)。
-菌群的生长速度系数(h-1)。
X-菌群的浓度(g〔菌群〕/L)。
YG-菌群的实际产量(g〔菌群〕/g〔培养基〕)。
qm-维持菌群生长对培养基消耗的系数(1/(g〔培养基〕.g〔菌群〕))。
对于一个只供给已经消毒的原料且状况稳定的恒化器来说,菌群的生长速度系数=D(D是培养基的稀释程度,是一个常数),则菌群在培养基中的浓度QS可以由D和菌群的产量来确定:
Qs=rs/X=D/Ys (2)
其中: YS-监测得到的菌群的产量(g〔菌群〕/g〔培养基〕)。
当只提供单一的碳源时,培养基将被细菌用于自身基本消耗和产生新的细菌。如果我们假设自身的消耗系数为0,即qm=0,所有的培养基都用于合成代谢产生新的细菌,这是理论上细菌产量最大的一种情况,在实际运行中,这个值将会比理论值小。为评价代谢过程的情况,qm 和 YG可以分别定量地做为对照而改变另一个量的值,以检测代谢过程进行的程度。将式(1)中的rS用式(2)中的代替并在两边同时除以DX,可以得到如下的方程式:
1/Ys=qm/D+1/YG (3)
由上式可以看出,只有qm 和 YG 保持在一定的浓度和速率上,使方程右边的值恒定不变,上面的假设才成立(即要使YS不变,方程式右边的两个变量在变化过程中,要保证方程值为常数),在一系列的监测数据中,以1/D为横坐标,以1/YS为纵坐标画图,将会得到一条直线,该直线的斜率为qm,截距为1/YG。
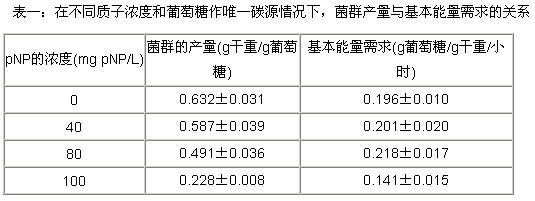 |
2 结论和讨论
2.1质子浓度对实验的影响
将恒化器中的培养基pH维持在7.0,以不同的稀释系列0,40,80和120 mg pNP/L 的pNP放入原料中,而菌群浓度和培养基的浓度随稀释多少而变化,此时会发现pNP的浓度会增加,且最大值出现在80-100 mg pNP/L。但是在120 mg pNP/L的时候我们会检测不到菌群的存在。在上述不同浓度段里,随着pNP浓度的增加,菌群的数据呈现出减少的趋势。同时发现,菌群培养基的消耗速率随pNP量的增加会呈增加趋势。Niejssel也由观测发现:在含有产气微生物的葡萄糖中加入二硝基酚和初始量的质子,碳源的消耗速度会增加。
尽管pNP存在可以降低反应器内菌群浓度和Dmax的值,但是这并不与上面提到的活性污泥工艺要实现泥水分离,污泥再回流到反应器的运行方案相矛盾,这两者是没有太大的相关性的。因此,菌群在曝气池内的浓度并不依赖于菌群的最大生长速率,而是依赖于回流污泥的量和污泥回流比。更有甚者,培养基的消耗速率随pNP的加入量增加而增加。在活性污泥工艺中,加大底物的更换速度来增加有机物的去除率这样的设想也是很合理的。对于已经建成的处理厂,我们可以增大反应器的负载的方法进行改造,对于新建的处理厂,可以减小反应器的有效容积,以获得更好的运行效果。
方程(3)告诉我们,1/Ys (因变量)相对 1/D(自变量)的线性函数,限定了不同质子浓度下培养基的状况(浓度等条件),菌群的基本能量需求和细菌真实产量由菌群的衰减程度决定。为此,在一定的线性范围内,菌群的产量和基本能量需求以及相对误差值可以由表一求得。与其它几位工作者(斯道汉姆、拜特海深和沃森)的研究一致,菌群在不同pNP浓度下,其基本能量需求近似是一个恒定的值,没有什么太大的变化。菌群的产量可以指示性地反应出菌群的产率。当pH控制在7.0,营养中供应100 mg pNP/L时,菌群的产量可以降低62%。这个结果表明,在尽量减少ATP含量的情况下,生物细胞会优先将能量用于提供自己最基本的能量需求,而不是用于合成代谢。
2.2pH值改变质子浓度对菌群产量的影响
活性污泥工艺一般运行在pH为6-8之间。酸性环境将会影响解偶联剂的活性。我们在恒化培养器中,改变pH的值来研究pNP浓度对菌群产量和对培养基更新速率的关系。在一系列不同稀释比例,质子浓度保持在100mg pNP/L恒定情况下,在pH分别为7.0、6.4或6. 2±0.1时,并且供给营养稳定不变,测得菌群、培养基浓度和pNP浓度都是相当稳定的。在不同pH值下,不添加pNP时,观测菌群基本能量需求和实际产量并没有明显的变化(如表二所示)。
在100 mg pNP/L时,降低培养基的pH值会降低菌群的产量,但是对底物更换速度没有明显的影响。观测发现,菌群产量随pH值的降低而降低。在更低的pNP浓度下(低于100 mg pNP/L),pH对培养基消耗速度的影响更加明显。生物基本能量需求和实际产量的关系列于表二中。质子通过氧化磷酸化的解偶联作用很明显地降低了YG的值,在低pH的环境里,对生物基本能量需求没有明显的影响,如果不加pNP时,基本上观测不到有什么变化。
表二:葡萄糖作唯一碳源,不加pNP和加浓度为100 mg pNP/L的pNP两种情况下,菌群实际产量和基本能量需求在不同pH值下的关系表
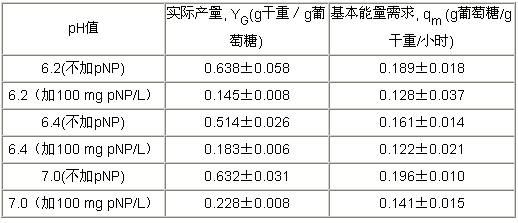 |
3 结论
1、在CFSTR反应器里,添加pNP可以降低生物的产量并且增加对有机物的消耗速率。
2、当用pNP刺激解偶联反应以降低细胞对能量的利用时,细胞会将能量优先用于自身最基本的需求,而不用于合成代谢产生新的细菌。
3、在进水中保持100 mg pNP/L 浓度的pNP,在pH为6.2的时候(和高pH值时相比较),可以大幅度地降低菌群产量并提高底物的消耗速率,这是因为pNP在氧化磷酸化过程中的解偶联作用使得能量都散失掉了,菌群只能通过降解更多的有机物来获得很少量的能量用于维持生命活动。
因此,解偶联剂的诱导作用可以在一定的范围内增加活性污泥工艺的处理效果并且可以降低污泥的产量。
4 文中引用介绍
1. 霍丹(1990年),奇切斯特人,威尔福德废水生物处理体系首席专家。
2. 霍斯登(1996年)。
3. 米切尔(1961年) ,研究化学渗透学说中解偶联作用在氧化磷酸化中电子和氢转移理论。摘自《自然》杂志191,144-148页。
4. 耐基斯尔(1997年),研究在有氧和培养基的恒化器中,2、4-二硝基酚对Klebsiella产气NCTC 418细菌生长的影响。《Fems Lett》1、47-50页。
5. 派特 (1975年),微生物和细胞培养方法的奠基人,牛津布莱克科学杂志出版物,66-74页。
6. 西蒙(1953年),研究二硝基酚毒性机理学,《Biol. Rev》28,453-479页。
7. 斯道汉姆和拜特海深 (1973年),研究在连续或间歇提供培养基情况下,生物对能量的利用情况。 《Biochim.Biophys.Acta》301,53-70页。
8. 沃森(1970年),研究在氯化钠对稳定生长的状况下的啤酒细菌新陈代谢的影响。 《J.Gen.Microbiol》64,91-99页
9. 赛尔和柯尔利(1984年),研究假单细菌对O-硝基酚和M-硝基酚的降解。《J. Agric. Food Chem.》32,238-242页。

使用微信“扫一扫”功能添加“谷腾环保网”





